生物化学_Electron Transport Chain & Oxidative Phosphorylation
20.Electron Transport Chain & Oxidative Phosphorylation
- basic
- 整体上将电子传递给O2,建立H+梯度并利用合成ATP
- A total of 3 ATP per NADH and 2 ATP per FADH2 may be produced through the processes of electron transport and oxidative phosphorylation
- Combining glycolysis and the TCA cycle gives
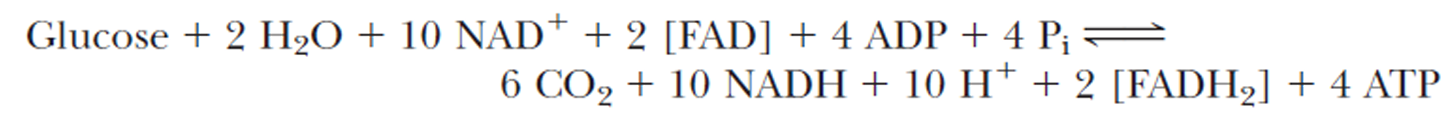
注意一个glycose进行一次glycolysis和两次TCA
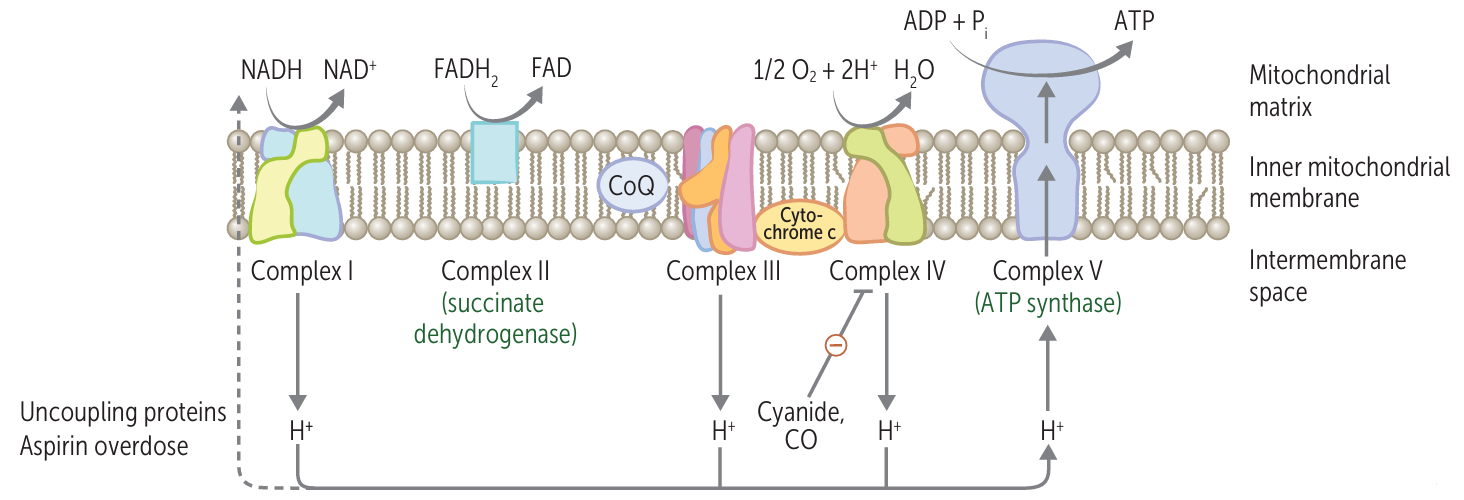
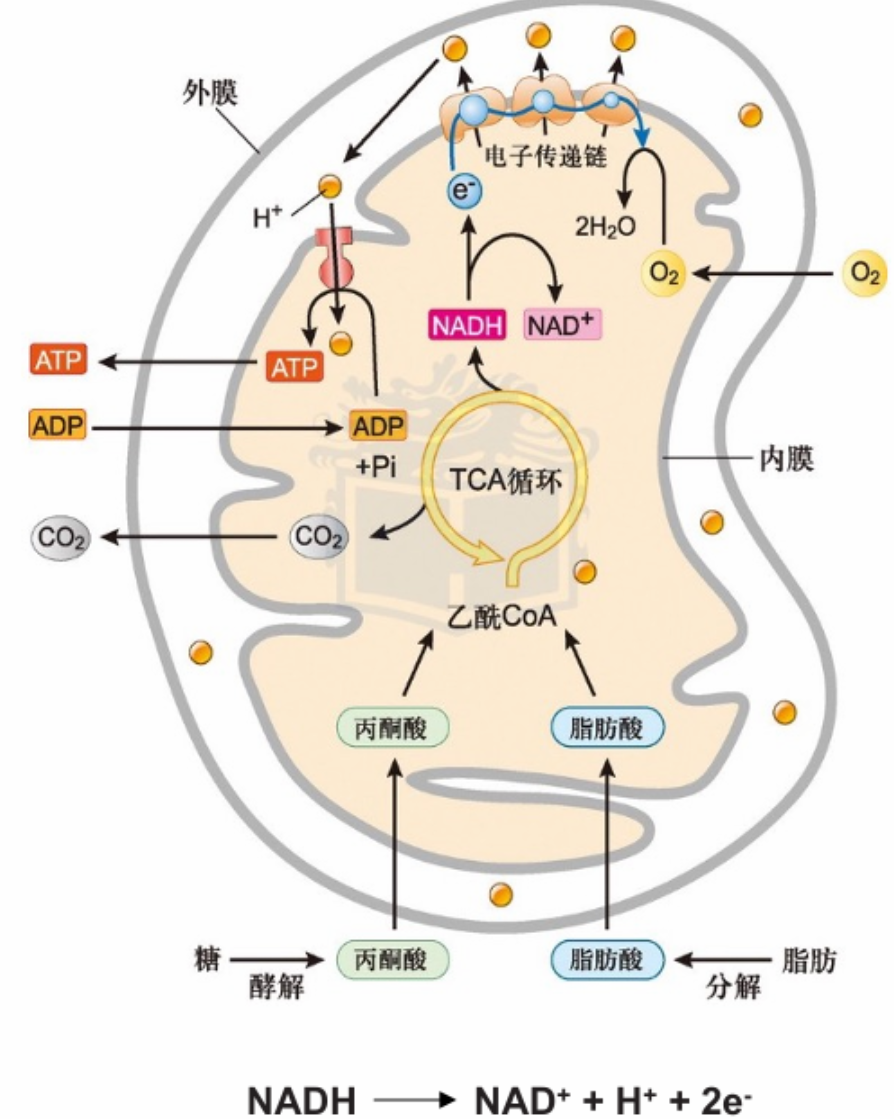
- Electrons stored in the form of the reduced coenzymes, NADH or [FADH2], are passed through a highly organized chain of proteins and coenzymes, the so-called electron transport chain, finally reaching O2(the terminal electron acceptor)
- This reoxidation process involves the removal of both protons and electrons from NADH and FADH2
- Each component of the chain can exist in (at least) two oxidation states, and as electrons move through the chain from NADH (or FADH2) to O2,each component is successively reduced and reoxidied.
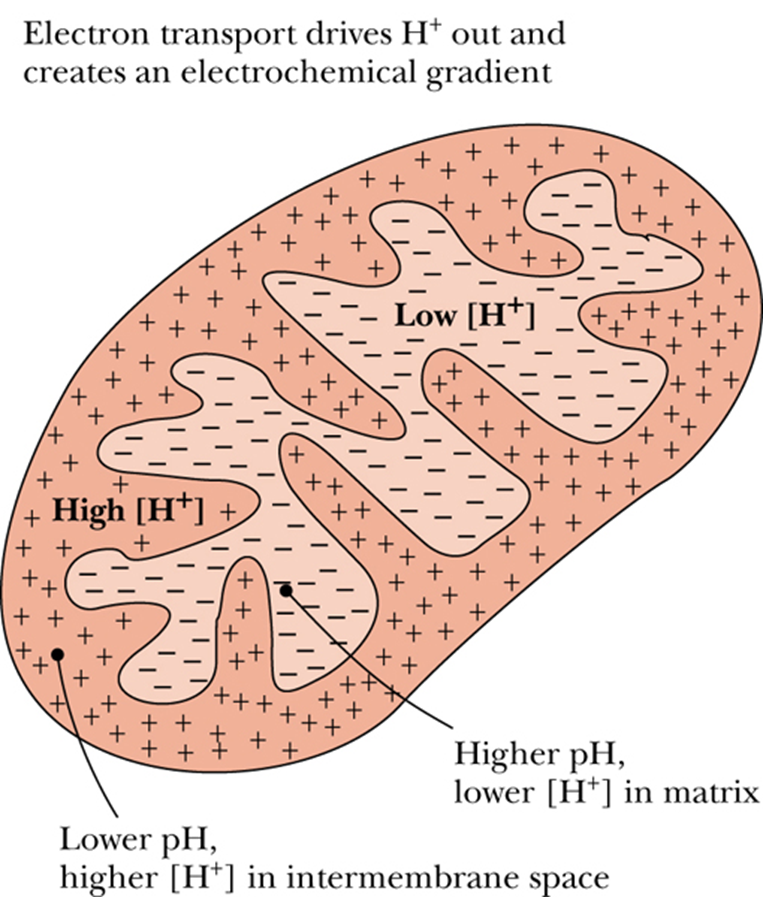
- In the course of electron transport, a proton gradient is established across the inner mitochondrial membrane, it is the energy of this proton gradient that drives ATP synthesis.
- 图
- 总结
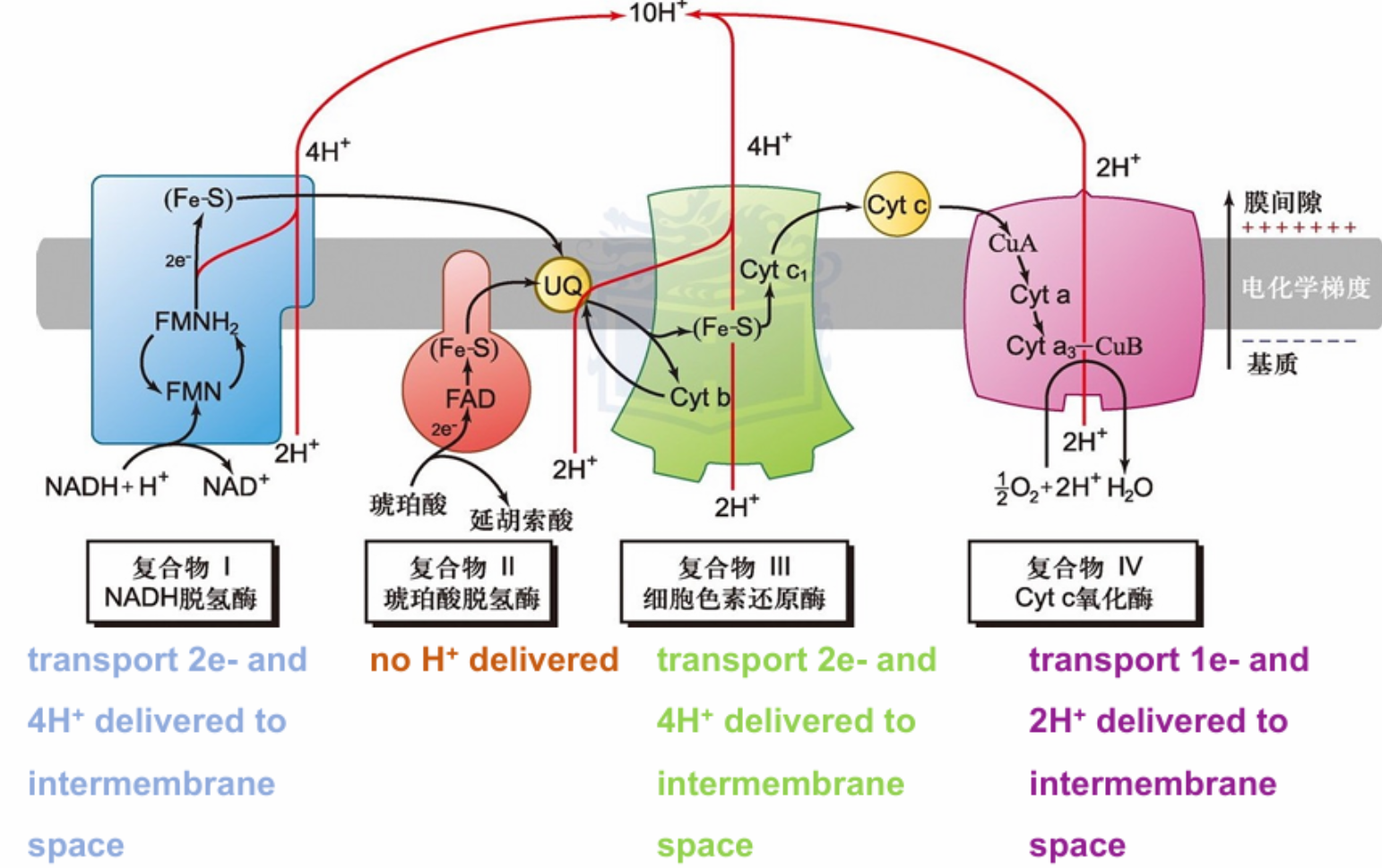
- NADH的2e-传递有10H+穿膜
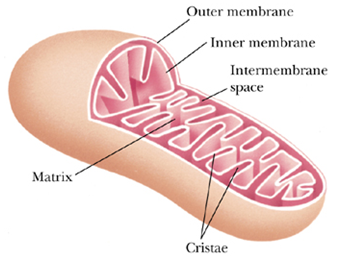
- 线粒体结构
- Mitochondria are surrounded by a simple outer membrane and a more complex inner membrane.
- outer
- the outer membrane functions mainly to maintain the shape of the mitochondrion
- contains significant amounts of porin孔蛋白
- a transmembrane protein, that forms large channels across the membrane
- permitting free diffusion of molecules with molecular weights of about 10,000 or less
- 图
- inner
- The inner membrane is richly packed with proteins and is extensively folded as cristae 嵴, provide the inner membrane with a large surface area in a small volume.
- outer
- The space between the inner and outer membranes is referred to as the intermembrane space.
- The space inside the inner mitochondrial membrane is called the matrix
- contains most of the enzymes of the TCA cycle and fatty acid oxidation
An exception, succinate dehydrogenase of the TCA cycle is located in the inner membrane.
- contains most of the enzymes of the TCA cycle and fatty acid oxidation
- 图
- In addition, mitochondria contain circular DNA molecules, ribosomes and the enzymes required to synthesize protein coded within the mitochondrial genome.
- The cytosolic side, where H accumulates, is referred to as the P (positive) face, the matrix side is the N (negative) face.
线粒体内膜, P face对着intermembrane space, Nface 对着 matrix
- Mitochondria are surrounded by a simple outer membrane and a more complex inner membrane.
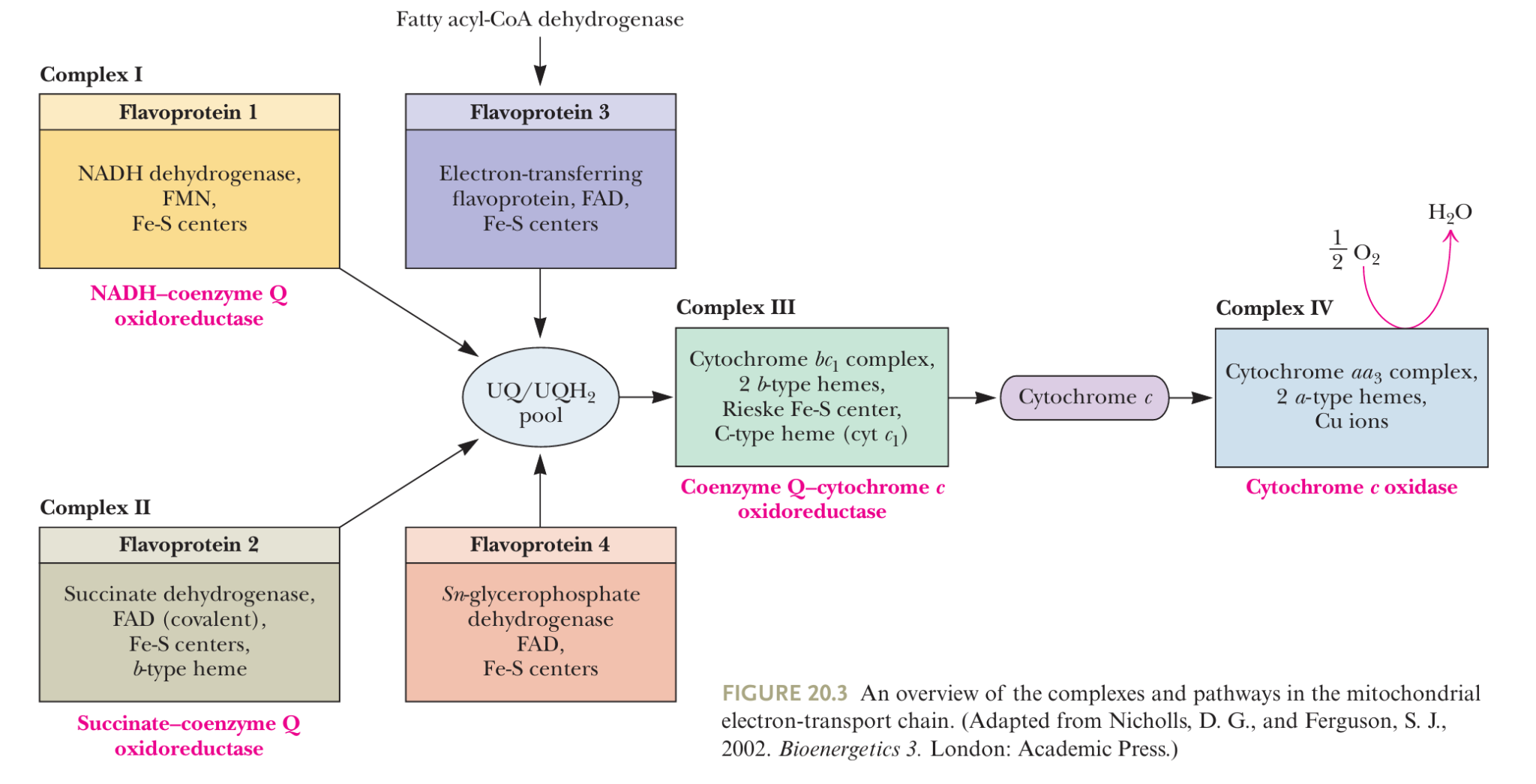
- molecular species
- (a) Flavoproteins 黄素蛋白
- which contain tightly bound flavin mono nucleotide (FMN) or FAD as prosthetic groups
- may participate in one- or two-electron transfer events.
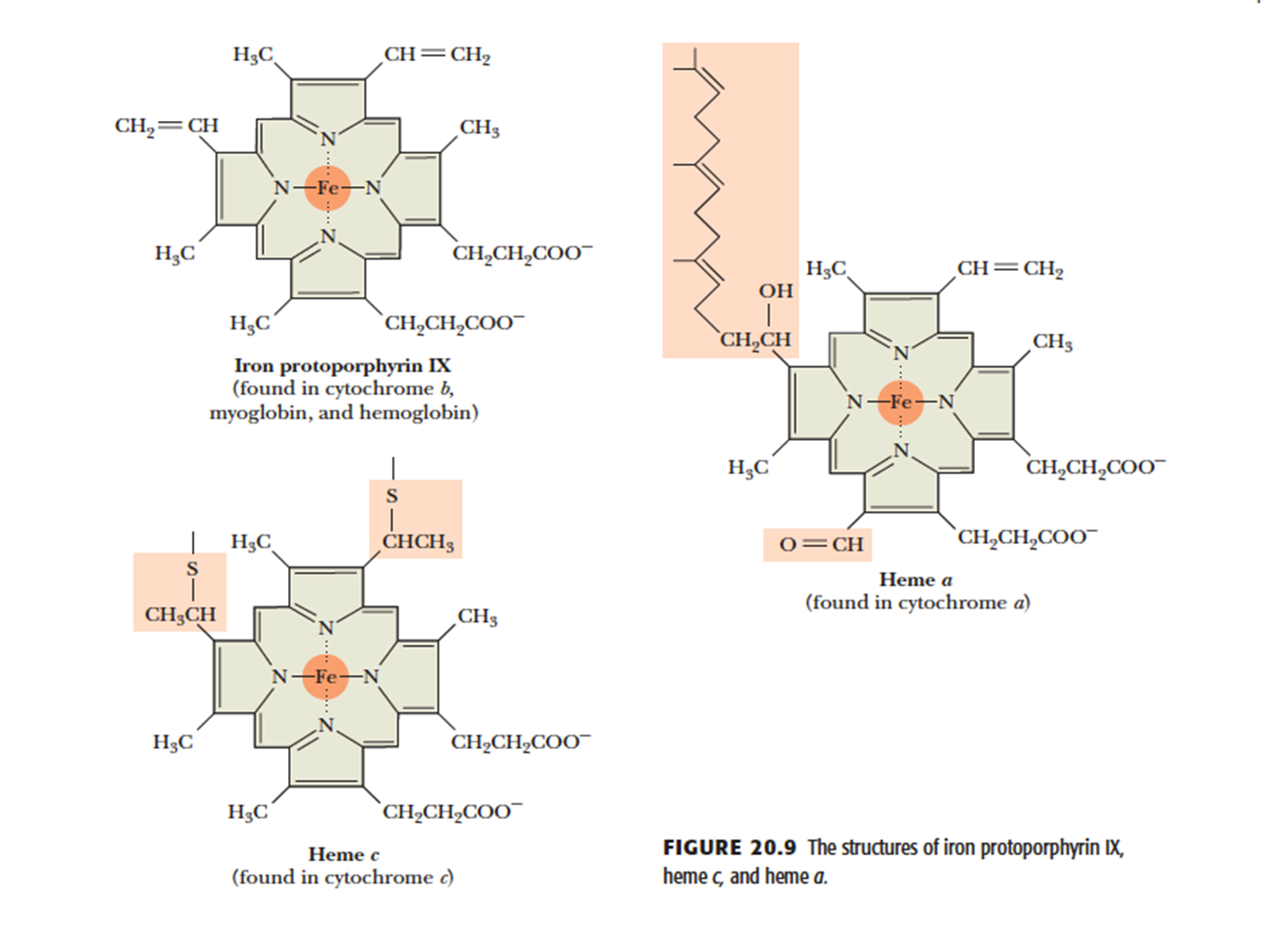
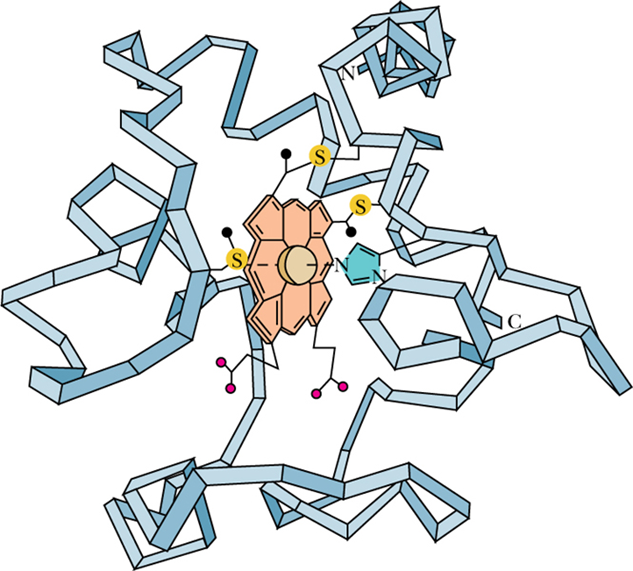
- (b) Cytochromes 细胞色素
- are proteins containing heme 亚铁血红素 prosthetic groups, which function by carrying or transferring electrons
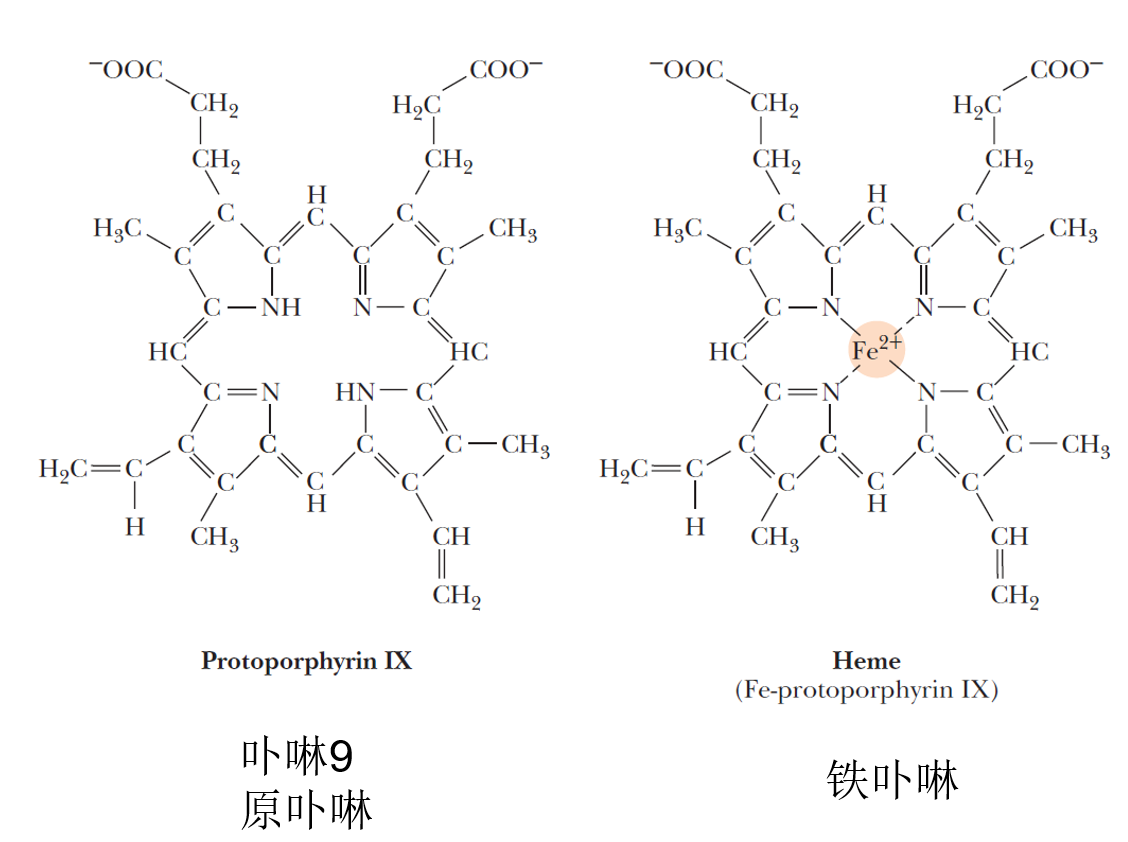
- heme
- Heme is formed when protoporphyrin IX binds Fe2+
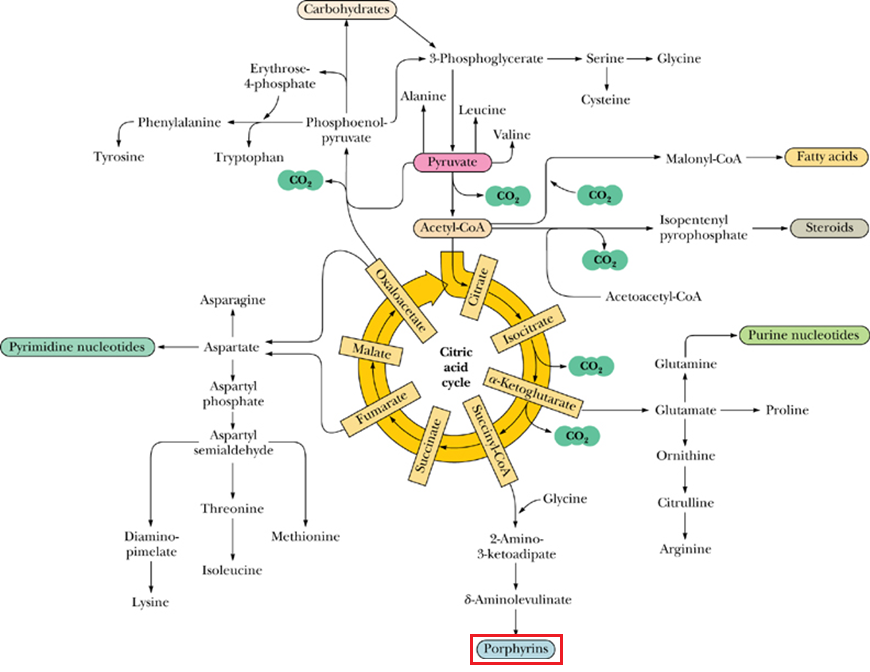
- The TCA cycle provides intermediates for many biosynthetic processes in the cell, 比如 porphyrin 卟啉 bu lin
- 图
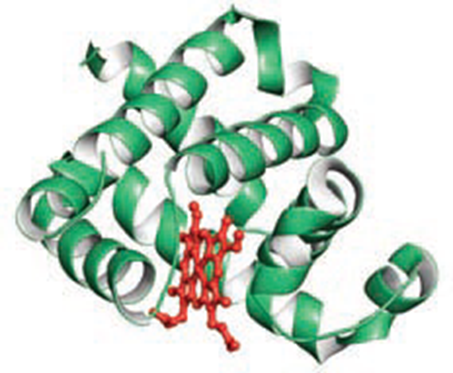
- Myoglobin (Mb) 肌红蛋白
- Oxygen binds to Mb via its heme
- Myoglobin is an oxygen-storage protein of muscle while Hemoglobin is the oxygen carrier
- Oxygen binds to Mb via its heme
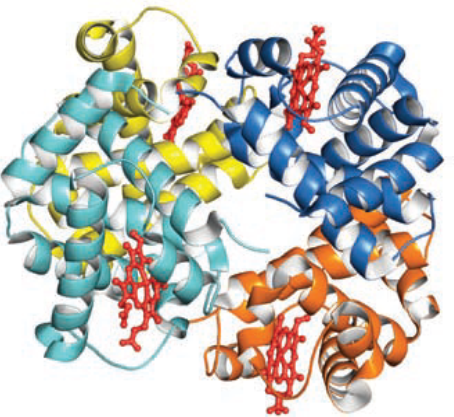
- Hemoglobin (Hb) 血红蛋白
- Hb is a tetramer.
- It consists of four polypeptide chains, each of which is very similar structurally to the Mb polypeptide chain, and each has a heme group.
- So, a Hb molecule can bind 4 O2 molecules
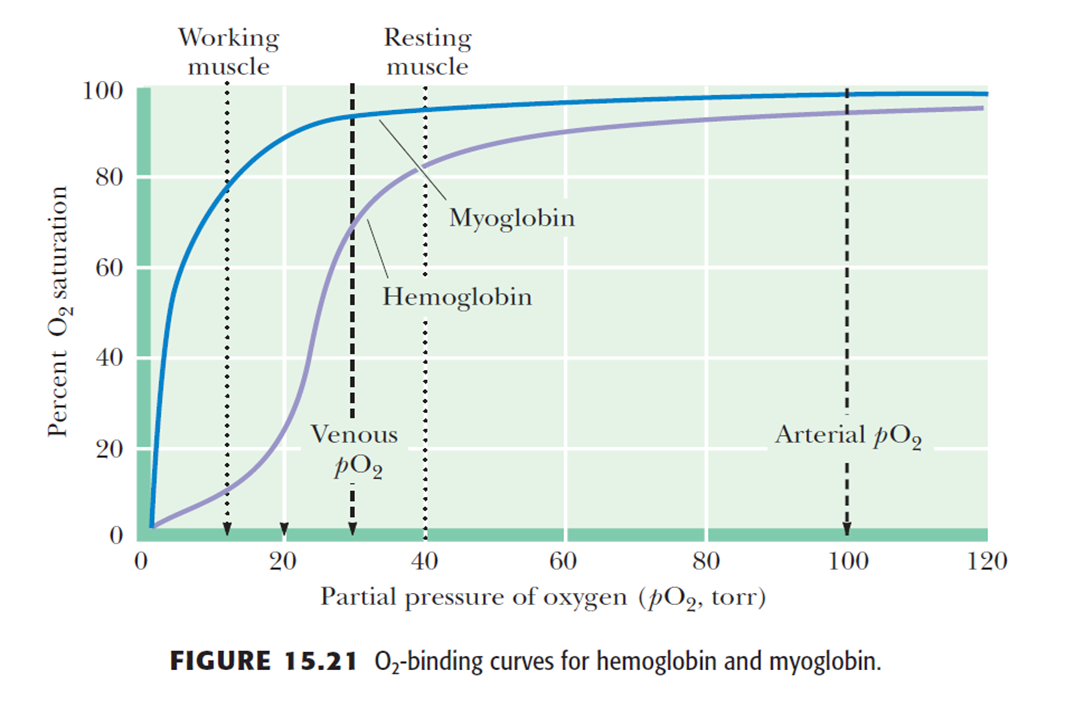
- O2 Binding Alters Hb Conformation. The binding of O2 to one heme of Hb makes it easier for the Hb molecule to bind additional equivalents of O2
- Hb is a tetramer.
- Hb displays sigmoid-shaped O2-binding curves. In contrast, Mb’s interaction with oxygen obeys classical Michaelis–Menten-type 米氏曲线 substrate saturation behavior.
- Heme is formed when protoporphyrin IX binds Fe2+
- Several cytochromes including cytochromes b, c, c1, a, and a3.
- Cytochromes are one-electron transfer agents, in which the heme iron is converted from Fe2+ (ferrous) to Fe3+ (ferric iron) and back.
- (c) iron–sulfur proteins 铁硫蛋白
- participate in one-electron transfers involving the Fe2+ and Fe3+ states
- Three types of proteins involved in this chain (a) (b) (c) possess electron-transferring prosthetic groups 辅基
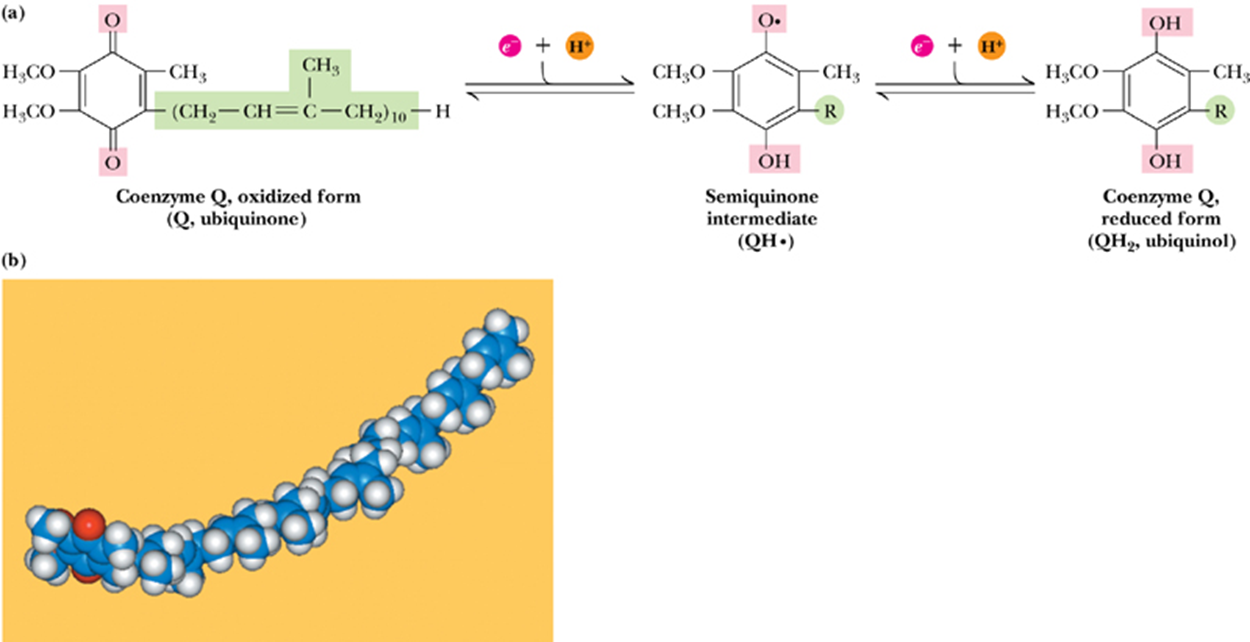
- (d) Coenzyme Q
- Q also called ubiquinone泛醌(and abbreviated CoQ or UQ)
- can function in either one or two-electron transfer reactions
- (e) Protein-bound copper
- a one-electron transfer site
- converts between Cu+ (cuprous) and Cu2+ (cupric)
- All these intermediates except for cytochrome c are membrane-associated
either in the mitochondrial inner membrane of eukaryotes or in the plasma membrane of prokaryotes
- (a) Flavoproteins 黄素蛋白
- 整体上将电子传递给O2,建立H+梯度并利用合成ATP
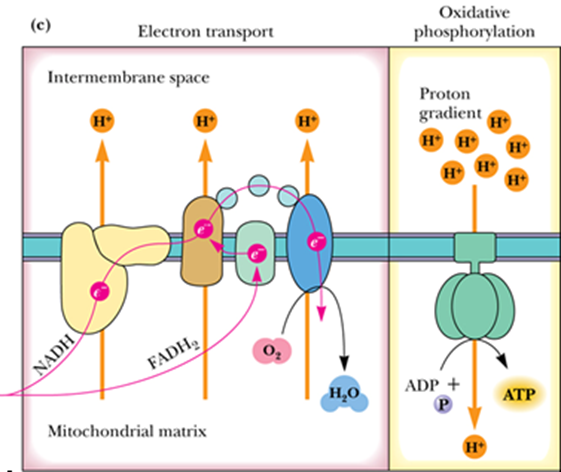
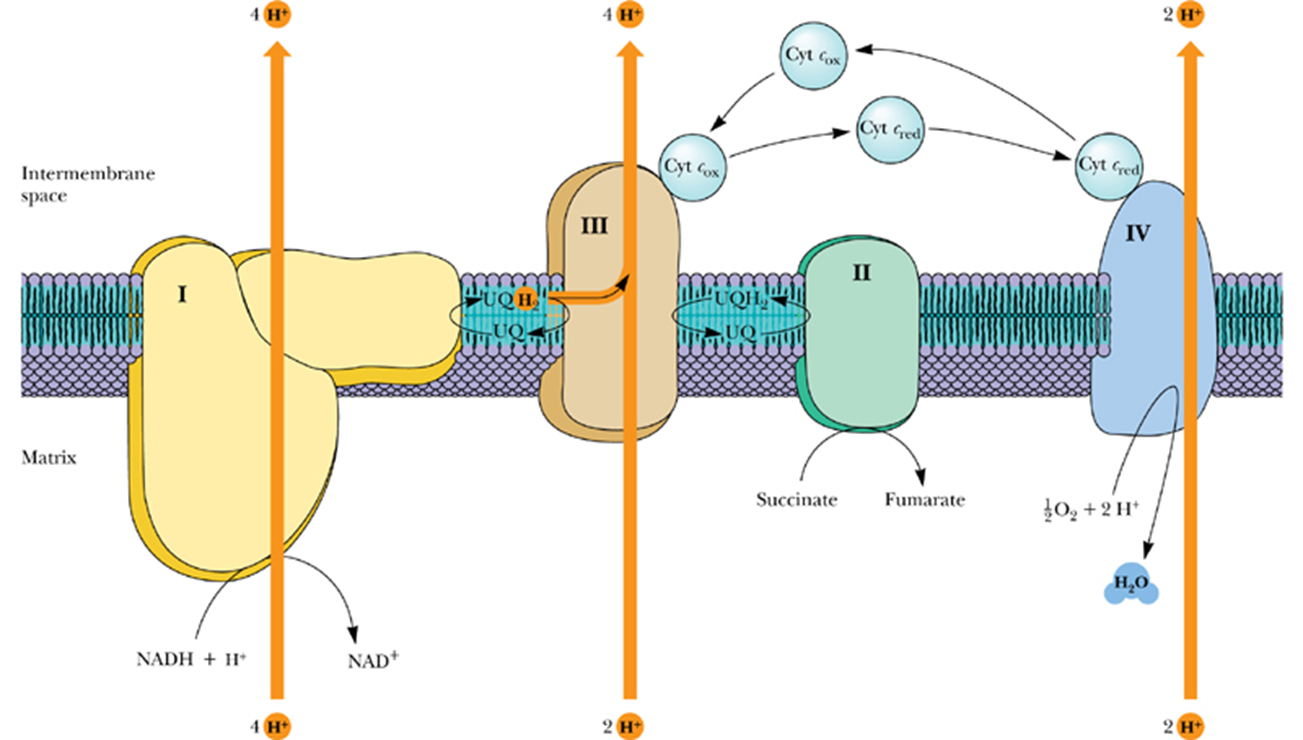
- Complex
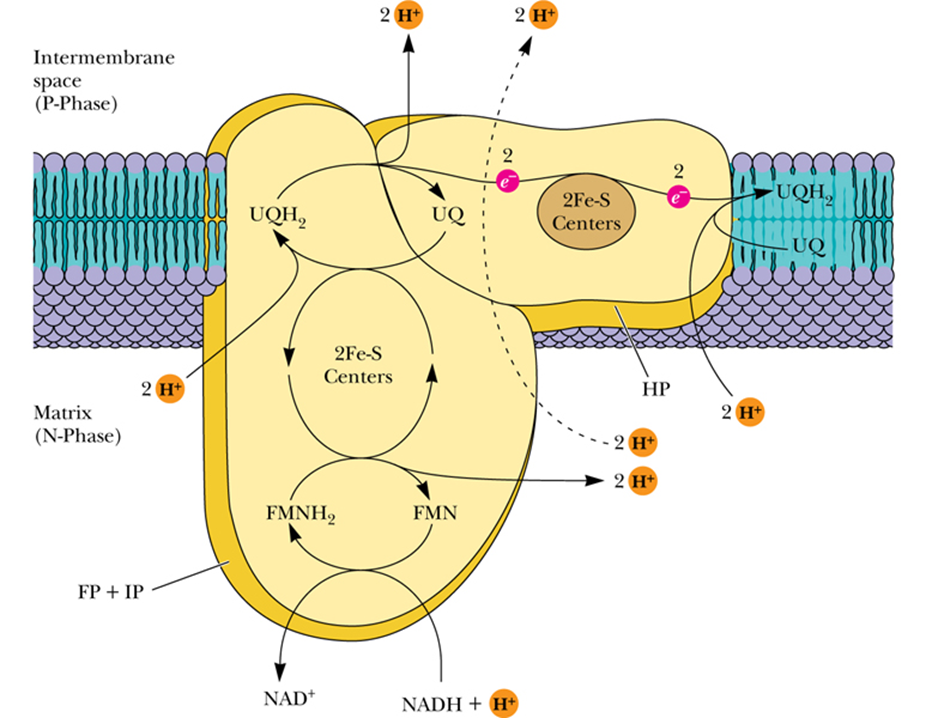
- Complex I - NADH–Coenzyme Q Reductase
NADH dehydrogenase- 图
- transfers a pair of electrons from NADH to coenzyme Q
- coenzyme Q
- a small, hydrophobic, yellow compound
- a mobile electron carrier
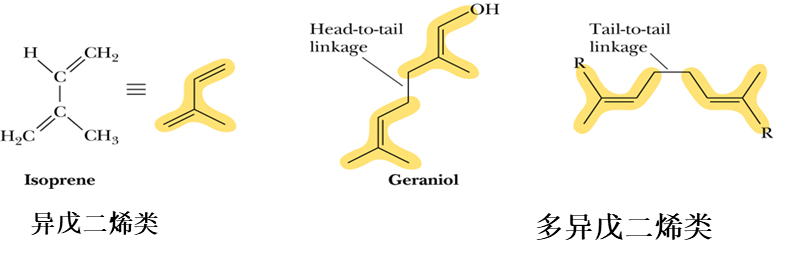
- Its isoprenoid 类异戊二烯 tail makes it highly hydrophobic, and it diffuses freely in the hydrophobic core ofthe inner mitochondrial membrane.
- As a result, it shuttles electrons from Complexes I and II to Complex III
- link between glycolysis, TCA cycle, fattyacid oxidation, and the electron transport chain
- 4 H+ transported per two electrons passed from NADH to UQ
- 图
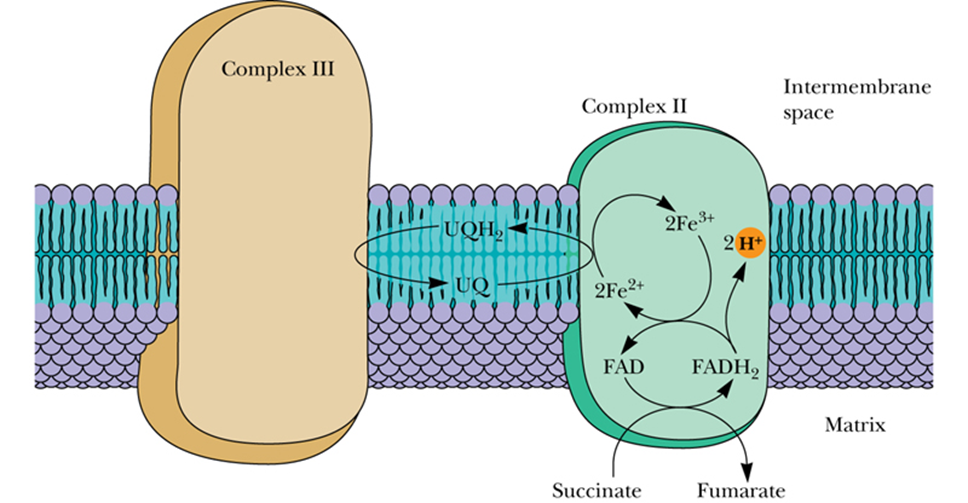
- Complex II - Succinate–Coenzyme Q Reductase
succinate dehydrogenase- 图
- The only TCA cycle enzyme that is an integral membrane protein in the inner mitochondrial membrane
- link between the TCA cycle and electron transport
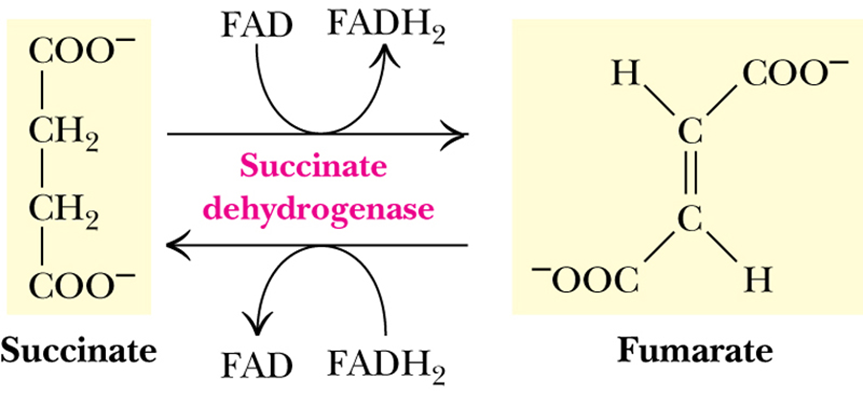
- When succinate is converted to fumarate in the TCA cycle, concomitant伴随着 reduction of bound FAD to FADH2 occurs in succinate dehydrogenase.
- This FADH2 transfers its electrons immediately to Fe-S centers, which pass them on to UQ.
- Electron flow from succinate to UQ
- Proton transport does not occur in this complex
- 图
- Complex III - Coenzyme Q–Cytochrome c Reductase
- 图
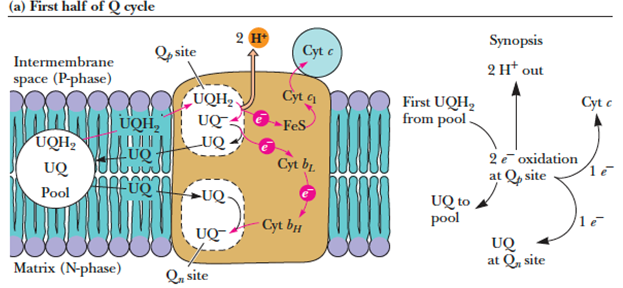
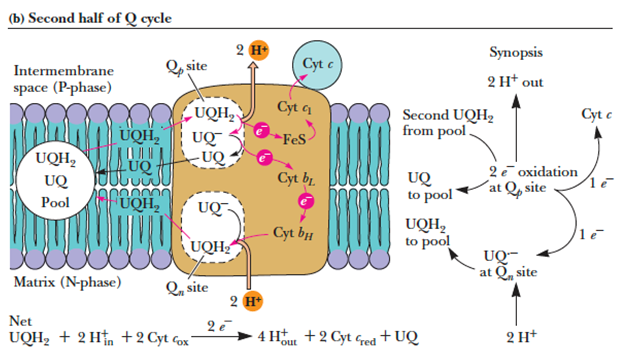
- In the third complex of the electron transport chain, reduced coenzyme Q (UQH2) passes its electrons to cytochrome c via a unique redox pathway known as the Q cycle.
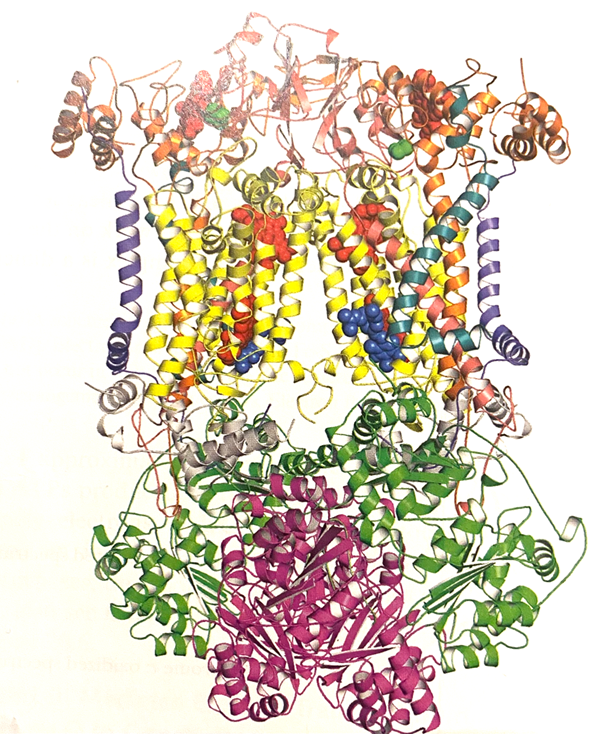
- structure
- The structure of UQ-cyt c reductase, as known as cytochrome bc1.
- The complex is dime
- the 𝛂-helices of cytochrome b (yellow) define the transmembrane domain of the protein.
- Heme groups are shown in red, Fe/S centers in green and associated phospholipids in blue.
- The bottom of the structure as shown extends approximately 75 Å into the mitochondrial matrix and the top of the structure as shown extends about 38 Å into the intermembrane space
- 图
- Cytochrome c
- Cytochrome c is the only one of the cytochromes that is water-soluble.
- It is globular, the planar heme group lies near the center of the protein, surrounded mainly by hydrophobic protein residues
- Cytochrome c, like UQ, is a mobile electron carrier.
- It associates loosely with the inner mitochondrial membrane to acquire electrons from Complex III,and then it migrates along the membrane surface and transfers electrons to complex IV
- Q cycle
- The electron-transfer pathway following oxidation of the first UQH2 at the Qp site near the cytosolic face of the inner membrane, releases 2 H+
- The pathway following oxidation of a second UQH2
- The electron-transfer pathway following oxidation of the first UQH2 at the Qp site near the cytosolic face of the inner membrane, releases 2 H+
- 图
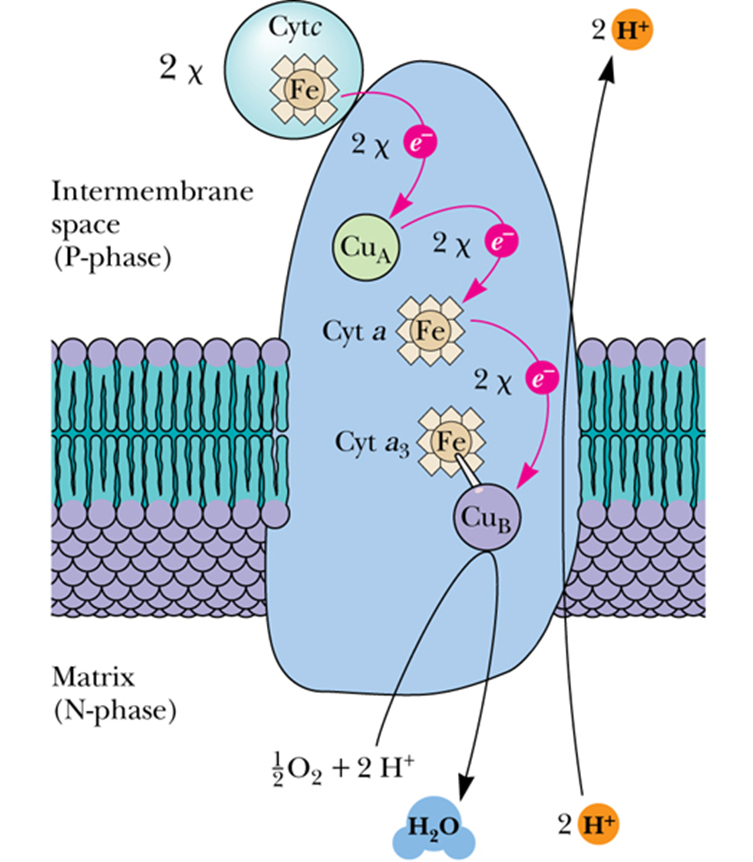
- Complex IV - Cytochrome c Oxidase
- 图
- accepts electrons from cytochrome c and directs them to the four-electron reduction of O2 to form H2O
- Electron transfer through Complex IV begins with binding of cytochrome c to the CuA center, next to heme a, and finally to the CuB/heme a3 active site, where O2 is reduced to H2O
- Transfer of 4 electrons through this complex drives the transport of 4 H+
- 图
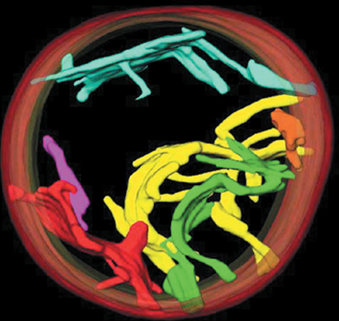
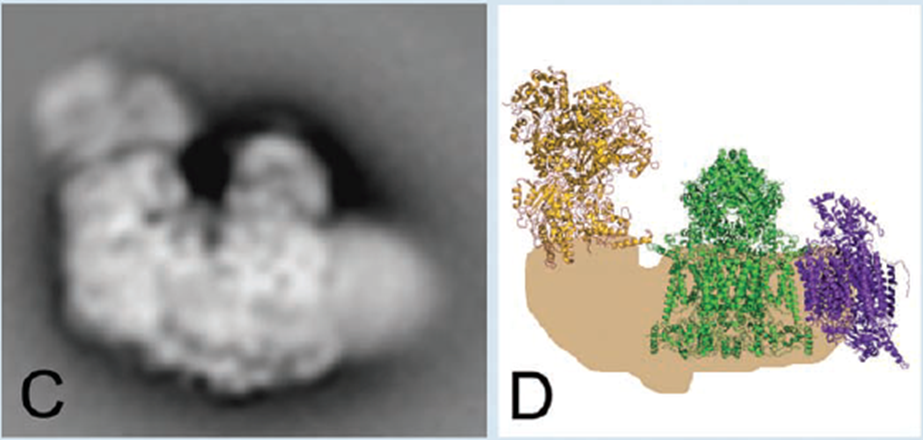
- The Complexes of Electron Transport May Function as Supercomplexes
- An electron microscopy image of a supercomplex formed from Complex I, Complex III,and Complex IV, and a model of this complex.
- 这些complex可能合在一起形成respirasomes呼吸体
- An electron microscopy image of a supercomplex formed from Complex I, Complex III,and Complex IV, and a model of this complex.
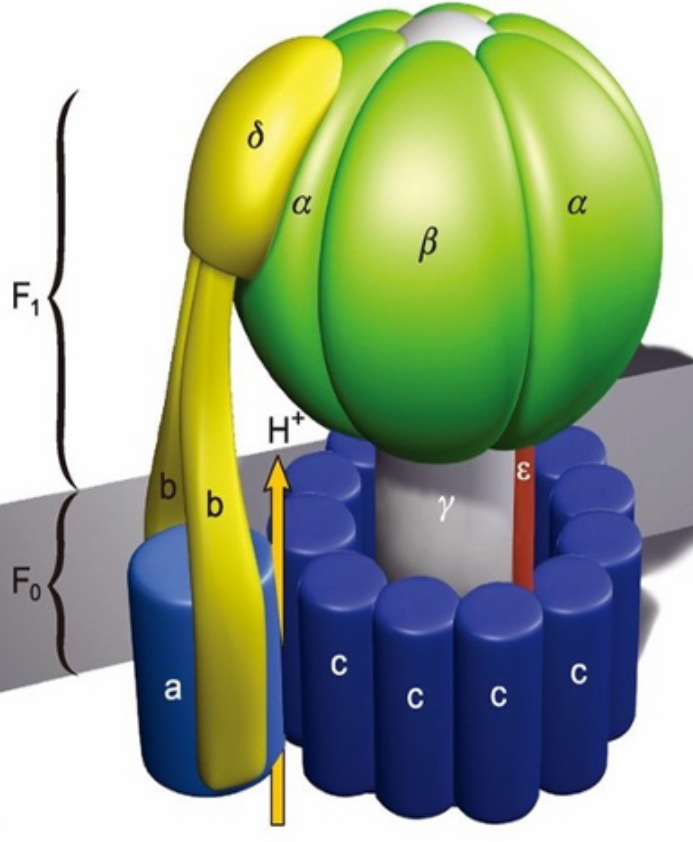
- Complex V - ATP Synthase
- The proton and electrochemical gradients across the inner mitochondrial membrane generated by electron transport represents a source of potential energy
- the ATP synthase projections凸起结构 appear on the matrix-facing surface of the inner membrane
- 上面对着matrix, 下面对着intermembrane space
- 图
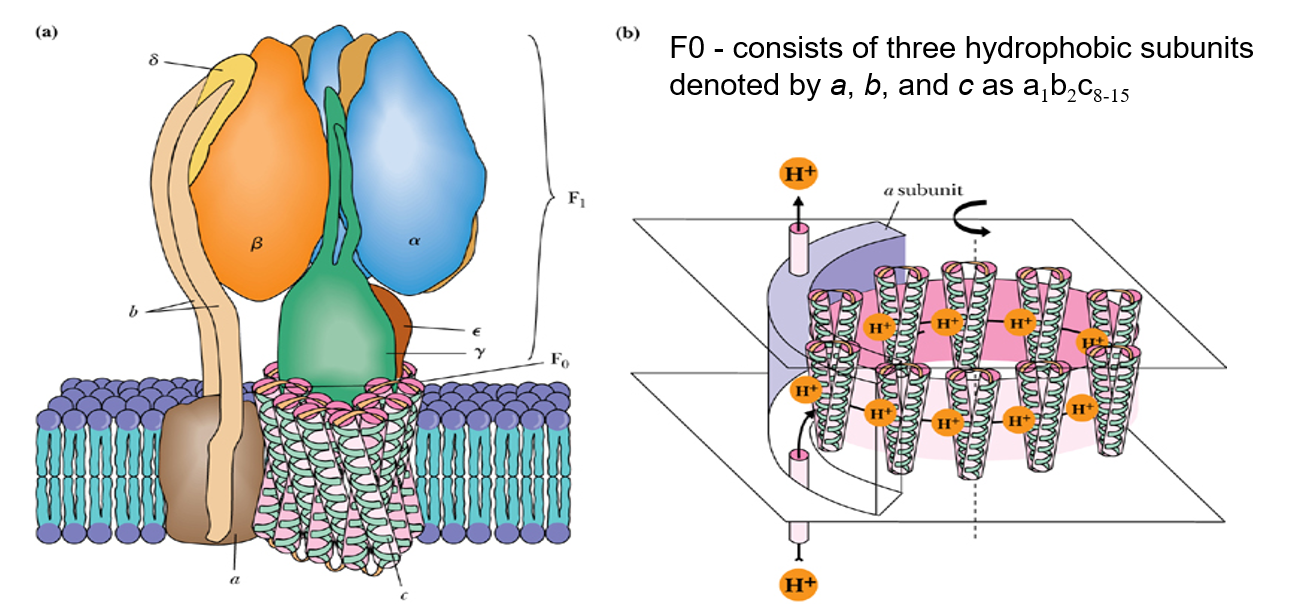
- Consists of Two Complexes
- F1
- hydrophilic
- The spheres observed make up the F1 unit, which catalyzes ATP synthesis
- F0
- hydrophobic
- integral membrane protein aggregate called the F0 unit.
- F0 forms the transmembrane pore or channel through which protons move to drive ATP synthesis
- c在不同生物中数量不同, 8-15个, 转一圈合成3ATP, 8-15/3 ≈ 3, 相当于3个H+穿过合成一个ATP
- F1
- Flow of protons through the structure turns the rotor and drives the cycle of conformational changes in α and β that synthesize ATP
- 只有底下这一圈c是在转的,其他都是固定的
- 用H+流动drive conformational change
- α和β都能结合ADP,只有α能催化ATP合成
- β 有3种conformation - Loose,Tight,Open
- rotor转子每转120°构象变化一次,合成一个ATP
- The proton and electrochemical gradients across the inner mitochondrial membrane generated by electron transport represents a source of potential energy
- Complex I - NADH–Coenzyme Q Reductase
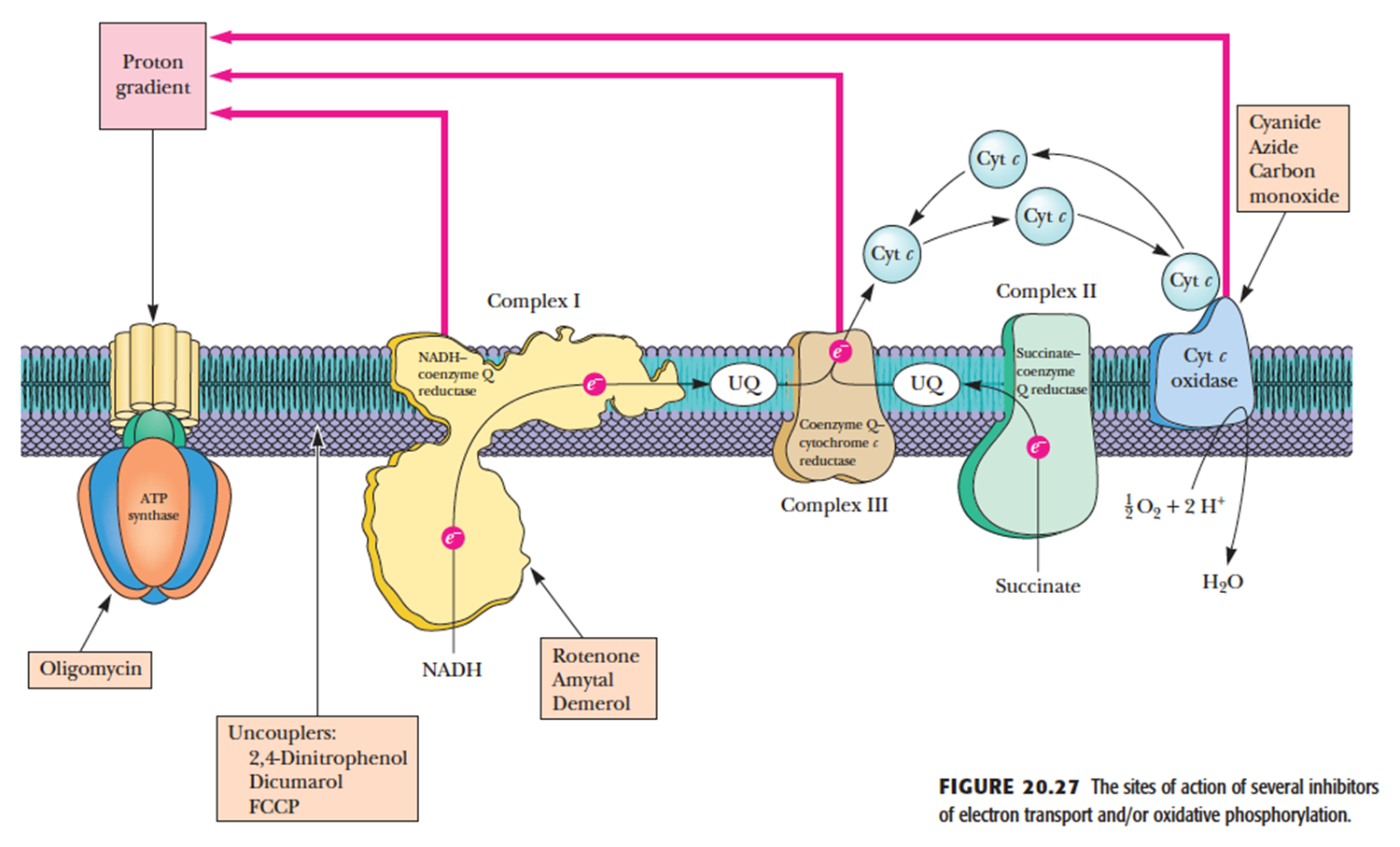
- Inhibitors
- Complexes I, II, and III Block Electron Transport Inhibitors
- Rotenone 鱼藤酮, 杀虫剂,抑制NADH-UQ
- Amytal 安密妥
- Demerol 杜冷丁
- Complex IV Inhibitors
- Cyanide 氰化物
- Azide 叠氮化物
- Carbon monoxide 一氧化碳
- ATP Synthase Inhibitors
- Oligomycin 寡霉素
- DCCD
- 图
- Complexes I, II, and III Block Electron Transport Inhibitors
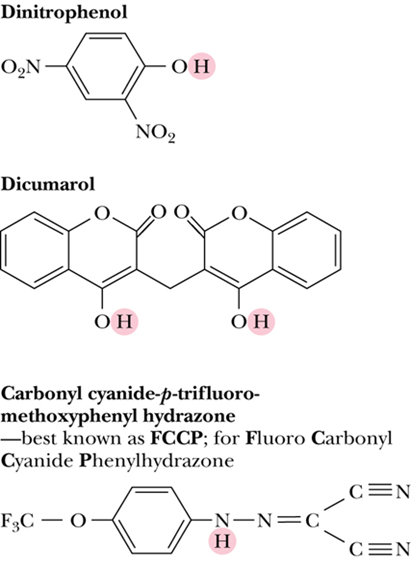
- Uncouplers 解偶联剂
- disrupt the tight coupling between electron transport and the ATP synthase
- Uncouplers act by dissipating the proton gradient across the inner mitochondrial membrane created by the electron transport system
- 不直接结合电子传递链或 ATP 合酶,但能破坏电子传递与 ATP 合成之间紧密偶联关系
- 他们会顺着质子梯度带质子穿过线粒体内膜, 把能量作为热量散失不合成ATP
- hydrophobic character and a dissociable proton 都具有疏水性和可以解离的质子
- 有一些耐寒动物, 冬眠动物, 新生动物, 植物会不合成ATP释放热量
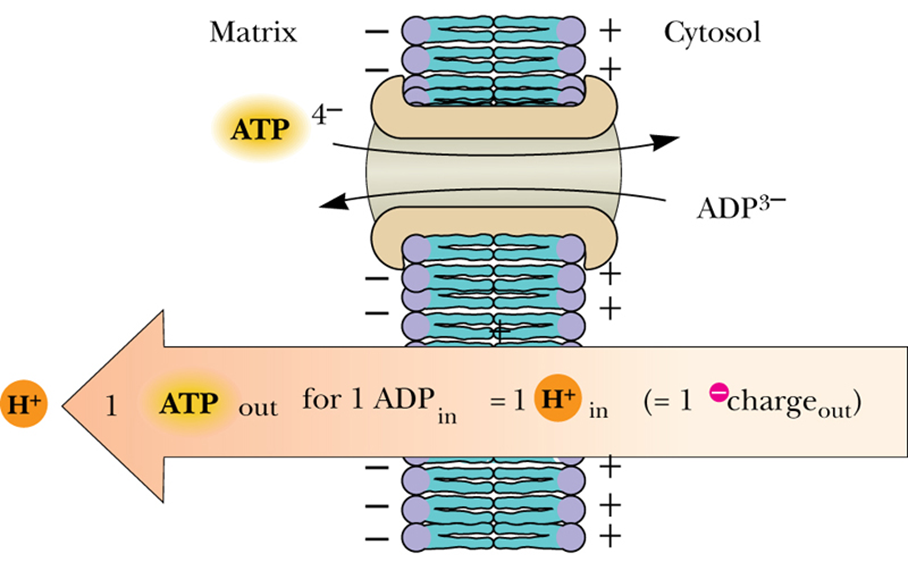
- ATP–ADP Translocase 移位酶
- ATP需要出线粒体发挥作用, ADP需要回线粒体
- highly charged ATP and ADP molecules do not readily cross biological membranes, 所以需要移位酶帮他们
- 1ATP出去 1ADP进来
- The translocase accounts for approximately 14% of the total mitochondrial membrane protein
- a homodimer同源二聚体 of 30-kD subunits
- It binds ATP on the matrix side, reorients to face the cytosol, and exchanges ATP for ADP, with subsequent movement back to the matrix face of the inner membrane
该蛋白先在基质侧结合 ATP,随后构象重排面向细胞质基质,将 ATP 与 ADP 进行交换,之后再重新转向线粒体内膜的基质侧,完成一次转运循环 - 正常生理情况下pH=7.2, ATP带-4电荷, ADP带-3电荷, 转运一次相当于1个H+进matrix
- 算上3H+合成一个ATP, 1H+让ATP出去, 相当于4H+的能量让一个ADP变成ATP, 1/4能量用于运输
- P/O ratio
- The P/O ratio is the number of molecules of ATP formed in oxidative phosphorylation per two electrons flowing through a defined segment of the electron transport chain
- 对于NADH, 2e-对应10H+出去, P/O = 10/4 = 2.5 ATP
- 对于FADH2, 2e-对应6H+出去, P/O = 6/4 =1.5 ATP
- These “consensus values” P/O ratios are of 2.7 and 1.6 for mitochondrial oxidation of NADH and [FADH2]. They may not reflect actual values and because these ratios may change depending on metabolic conditions, these estimates of ATP yield from glucose oxidation are approximate.
目前公认的 NADH 和 FADH₂线粒体氧化的 P/O 比值分别为 2.7 和 1.6, 但这些数值并非绝对, 由于代谢条件变化可能影响该比值,因此葡萄糖氧化过程中 ATP 产量的估算值仅为近似值 - All six carbons of glucose are liberated as CO2, and a total of four molecules of ATP are formed thus far in substrate-level phosphorylations. A total of 2.7 ATP per NADH and 1.6 ATP per FADH2 may be produced through the processes of electron transport and oxidative phosphorylation. Thus the 12 reduced coenzymes produced up to this point can eventually produce a maximum of 30.2 molecules of ATP in the electron transport and oxidative phosphorylation pathways. Thus, a glucose can produce 34.2 ATP.
- 葡萄糖的 6 个碳原子均以 CO₂形式释放,截至目前(糖酵解 + 三羧酸循环阶段)通过底物水平磷酸化共生成 4 分子 ATP。通过电子传递链与氧化磷酸化过程,每分子 NADH 可产生约 2.7 分子 ATP,每分子 FADH₂可产生约 1.6 分子 ATP。因此,至此阶段生成的 12 分子还原型辅酶(NADH+FADH₂),最终可通过电子传递与氧化磷酸化途径最多生成 30.2 分子 ATP. 综上,1 分子葡萄糖完全氧化可产生 34.2 分子 ATP
- shuttle system
让细胞质中糖酵解产生的NADH参与电子传递- Eukaryotic cells have a number of shuttle systems that harvest the electrons of cytosolic NADH for delivery to mitochondria without actually transporting NADH across the inner membrane
- Glycerophosphate Shuttle
- 3-磷酸甘油穿梭途径
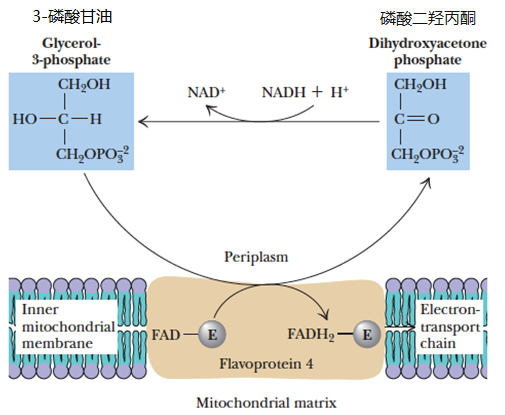
Glycerophosphate is different from glyceraldehyde-3-phosphate (G-3-P)3-磷酸甘油醛
- The glycerophosphate shuttle couples the cytosolic oxidation of NADH with mitochondrial reduction of [FAD]
- Cytosolic NADH oxidized via this shuttle route yields only 1.6 molecules of ATP. The cell “pays” with a potential ATP molecule for the convenience of getting cytosolic NADH into the mitochondria.
通过该穿梭途径氧化的胞质 NADH 仅产生 1.6 分子 ATP, 细胞为了将胞质 NADH “送入” 线粒体,需 “付出” 1 个潜在的 ATP 分子作为代价 - The glycerophosphate shuttle is essentially irreversible, and even when NADH levels are very low relative to NAD, the cycle works effectively
3-磷酸甘油穿梭系统本质上是不可逆的,即使 NADH 相对于 NAD 的水平很低,该循环仍能有效运转
- 3-磷酸甘油穿梭途径
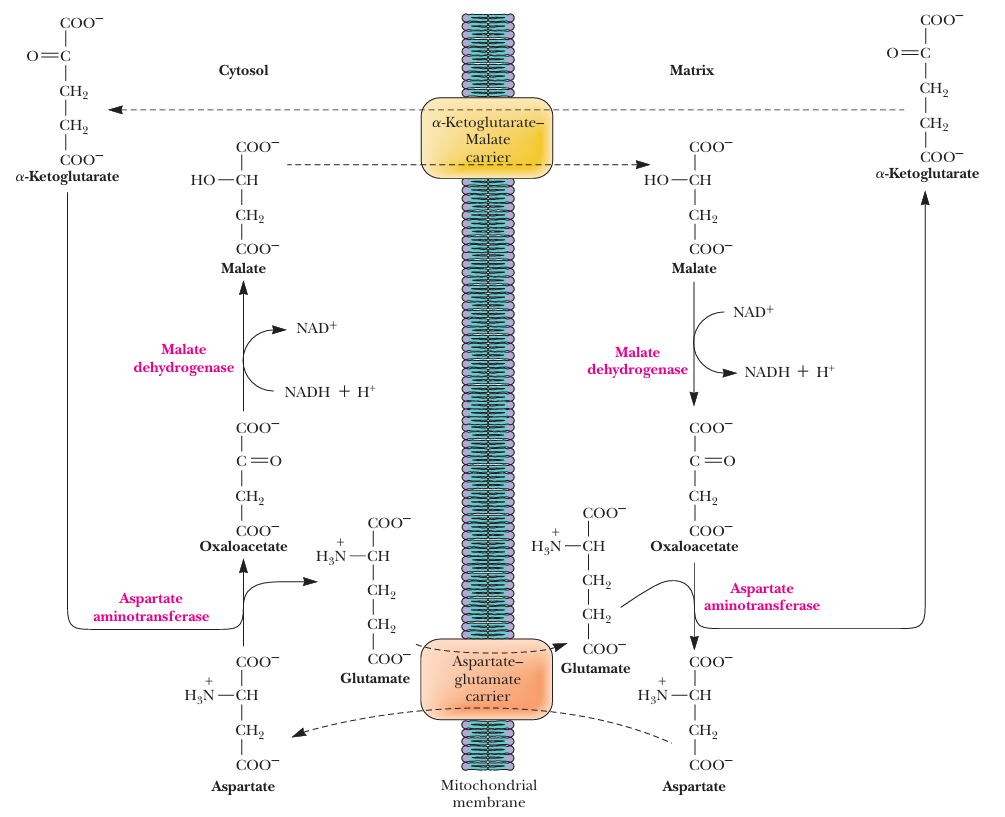
- The Malate–Aspartate Shuttle
- 苹果酸-天冬氨酸途径
- Oxaloacetate草酰乙酸 is reduced in the cytosol, acquiring the electrons of NADH and to generate Malate苹果酸
- Malate苹果酸 is transported across the inner membrane, converting NAD to NADH in the matrix and to generate oxaloacetate草酰乙酸
- The oxaloacetate草酰乙酸 can not cross the inner membrane, and must be transaminated to form aspartate天冬氨酸
- reversible
- it works only if the NADH/NAD ratio in the cytosol is higher than the ratio in the matrix.
- Because this shuttle produces NADH in the matrix, the full 2.7 ATPs per NADH are recovered
- 苹果酸-天冬氨酸途径
- The Net Yield of ATP from Glucose Oxidation Depends on the Shuttle Used
- 32 to 34.2 molecules of ATP permolecule of glucose
- Using the glycerol phosphate shuttle, is 32 ATP
- Using the malate–aspartate shuttle, is 34.2 ATP
- Prokaryotic cells(bacteria) need not carry out ATP/ADP exchange, potential to produce 38 ATP
细菌的电子传递链在细胞膜上
- 线粒体总结
- 哺乳动物细胞含有 800 至 2500 个线粒体
- 其他类型细胞的线粒体数量差异较大,少则 1-2 个,多则可达 50 万个
- 人类红细胞的功能仅为向组织运输氧气,因此完全不含线粒体
- 典型线粒体的直径约为 0.3-0.5 微米,长度为 0.5 微米至数微米
- over all shape is sensitive to metabolic conditions in the cell
- 从葡萄糖将能量转化为ATP的效率约为54%
- 图
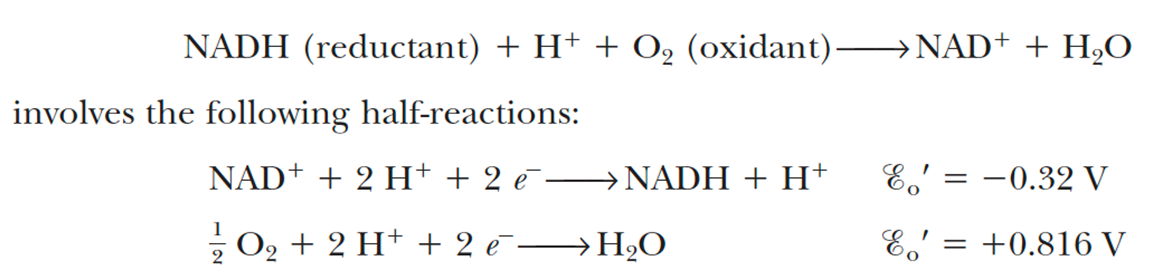
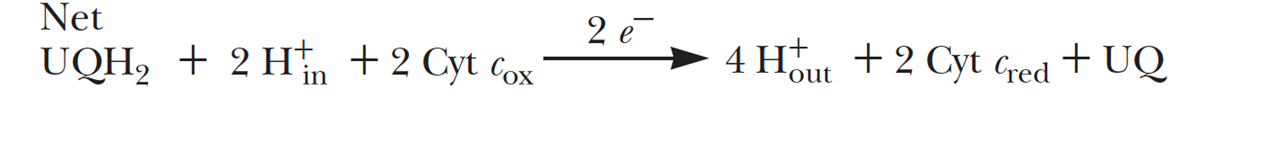
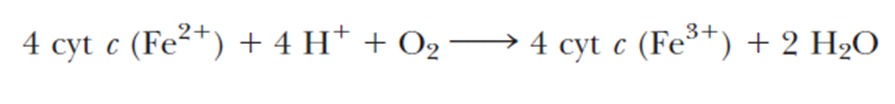
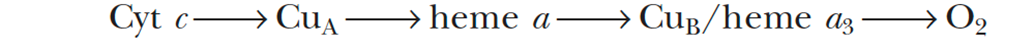

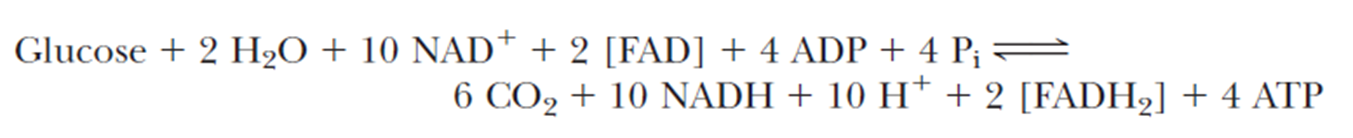
All articles in this blog are licensed under CC BY-NC-SA 4.0 unless stating additionally.
